Médico experto del artículo.

Nuevos artículos
Modelos experimentales de artrosis
Último revisado: 07.07.2025

Todo el contenido de iLive se revisa médicamente o se verifica para asegurar la mayor precisión posible.
Tenemos pautas de abastecimiento estrictas y solo estamos vinculados a sitios de medios acreditados, instituciones de investigación académica y, siempre que sea posible, estudios con revisión médica. Tenga en cuenta que los números entre paréntesis ([1], [2], etc.) son enlaces a estos estudios en los que se puede hacer clic.
Si considera que alguno de nuestros contenidos es incorrecto, está desactualizado o es cuestionable, selecciónelo y presione Ctrl + Intro.
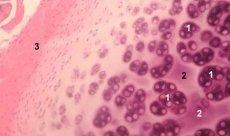
El cartílago es un tejido altamente especializado que contiene un solo tipo de células (condrocitos) y se caracteriza por la ausencia de vasos sanguíneos y linfáticos. El cartílago se nutre principalmente mediante la absorción del líquido sinovial. El metabolismo de los condrocitos está regulado por diversos factores solubles producidos localmente por los condrocitos y los tejidos circundantes. La función de los condrocitos también depende de la composición del entorno extracelular (tensión de oxígeno, concentración iónica, pH, etc.), la composición de la matriz extracelular (ECM), la interacción de las células con la matriz, y las señales físicas. El objetivo principal del modelado experimental es crear cultivos en el entorno extracelular sin modificar el fenotipo de las células maduras. El segundo objetivo es crear cultivos para estudiar la respuesta prematura, retardada, a corto plazo o prolongada de los condrocitos a señales químicas o físicas. Los estudios in vitro también ofrecen la oportunidad de estudiar el comportamiento de los condrocitos en la osteoartrosis. El tercer objetivo es desarrollar sistemas de cocultivo que permitan estudiar las interacciones de diversos tejidos en la articulación. La cuarta tarea es la preparación de implantes de cartílago para su posterior trasplante. Finalmente, la quinta tarea es el estudio de factores de crecimiento, citocinas o agentes terapéuticos capaces de estimular la reparación o inhibir la resorción del cartílago.
En las últimas décadas, se han creado varios modelos de cultivos celulares de cartílago articular, incluyendo cultivos en monocapa, cultivos suspendidos, cultivos de condrones, explantos, cocultivos y cultivos de células inmortales. Cada cultivo tiene sus propias ventajas y desventajas, y cada uno es adecuado para estudiar un aspecto específico del metabolismo de los condrocitos. Por lo tanto, los explantos de cartílago son un excelente modelo para estudiar el recambio de elementos de la matriz, que requiere receptores genuinos de la superficie celular e interacciones normales célula-matriz y matriz-célula. Al mismo tiempo, se recomienda estudiar los depósitos de matriz o los mecanismos que regulan el metabolismo de los condrocitos en un cultivo de células aisladas. Un cultivo en monocapa de baja densidad es necesario para estudiar el proceso de diferenciación celular. Los cultivos suspendidos en una matriz natural o sintética son un modelo para analizar la respuesta adaptativa de los condrocitos al estrés mecánico.
Cultivos de condrocitos
Se deben tener en cuenta varios puntos importantes al seleccionar tejido cartilaginoso para estudios in vitro. La composición de la matriz y la actividad metabólica de los condrocitos varían entre articulaciones, y esta última también depende de la profundidad de la ubicación de los condrocitos en el tejido. Estos datos se obtuvieron en varios experimentos en los que se estudiaron subpoblaciones aisladas de condrocitos de zonas de cartílago de diferentes profundidades. Se encontraron varias diferencias morfológicas y bioquímicas entre los condrocitos cultivados ubicados en las capas superficiales y profundas del cartílago articular. Las células superficiales sintetizan una matriz fibrilar escasa y pobre en proteoglicanos, mientras que las células más profundas producen una matriz rica en fibrillas y proteoglicanos. Además, las células superficiales producen relativamente más proteoglicanos pequeños no agregados y ácido hialurónico, y relativamente menos agrecano y queratán sulfato que los condrocitos más profundos. Otra característica distintiva importante del metabolismo de los condrocitos aislados de zonas de cartílago de diferentes profundidades es la respuesta a un estímulo exógeno. Según M. Aydelotte et al., los condrocitos bovinos de la zona superficial del cartílago fueron más sensibles a IL-1 que las células de la zona profunda.
El comportamiento celular también depende de la ubicación del tejido. Los condrocitos del cartílago de las costillas y de la oreja del mismo animal responden de manera diferente a factores de crecimiento como el factor de crecimiento de fibroblastos (FGF) y el TGF-beta. El FGF aumentó la incorporación de timidina, prolina y leucina en los condrocitos cultivados de las costillas, pero no en los de las orejas. El TGF-beta aumentó la incorporación de timidina en los condrocitos de las costillas y del cartílago de la oreja, pero no tuvo efecto sobre la incorporación de timidina y prolina en los condrocitos de la oreja. Las células del cartílago de las áreas de alto estrés difieren de las de las áreas de bajo estrés en el cartílago. Por lo tanto, los condrocitos del cartílago maduro de la articulación de la rodilla de oveja de la región central de la superficie articular de la tibia no cubierta por el menisco, que soporta la mayor carga in vivo, sintetizan menos agrecano, pero más decorina que las células de las zonas cubiertas por el menisco. Los autores también enfatizan la importancia de utilizar cartílago de zonas articulares idénticas al estudiar la función sintética de las articulaciones.
El metabolismo de los condrocitos y su respuesta a factores reguladores también dependen significativamente de la edad del donante, su desarrollo esquelético y la condición de las articulaciones de las que se extraen las células. En condrocitos humanos, se observa una disminución significativa en la respuesta proliferativa con la edad. La mayor disminución se observa en donantes de 40 a 50 años y mayores de 60 años. Además, la gravedad de la respuesta proliferativa a factores de crecimiento (p. ej., FGF y TGF-beta) disminuye con el envejecimiento. Además de los cambios cuantitativos en la proliferación de condrocitos, también hay cambios cualitativos. Las células de donantes jóvenes (10-20 años) responden mejor al factor de crecimiento derivado de plaquetas (PDGF) que al TGF-beta, mientras que se observa lo contrario en células de donantes adultos. Se utilizan varios mecanismos para explicar los cambios dependientes de la edad en la función sintética de los condrocitos y su respuesta a los factores de crecimiento. Estos incluyen una disminución en el número y la afinidad de los receptores de las células de superficie, cambios en la síntesis y bioactividad de los factores de crecimiento y las citocinas, y modificación de las señales post-receptor.
La condición patológica de las articulaciones también cambia la morfología y la actividad metabólica de los condrocitos. Así, J. Kouri et al. (1996) identificaron tres subpoblaciones de condrocitos en el cartílago en la osteoartrosis. Los condrocitos de la parte superficial y superior de la parte media del cartílago forman grupos y sintetizan una mayor cantidad de proteoglicanos y colágeno. El TGF-beta y el factor de crecimiento similar a la insulina (IGF) son capaces de estimular la síntesis de proteoglicanos por los condrocitos y neutralizar parcialmente los efectos de IL-1 y TNF-a. Los explantos de cartílago afectado por osteoartrosis y los condrocitos aislados del cartílago de un paciente con osteoartrosis son más sensibles a la estimulación del TGF-beta que los condrocitos del cartílago sano. Estas diferencias están muy probablemente asociadas con cambios fenotípicos en los condrocitos en las capas superiores del cartílago articular.
El aislamiento de condrocitos individuales se logra mediante el tratamiento secuencial de la matriz extracelular (ECM) con enzimas proteolíticas. Tras su liberación de la ECM, las células aisladas son ideales para estudiar la síntesis de novo de componentes de la matriz. Algunos autores utilizan únicamente colagenasa clostridial, mientras que otros preincuban el cartílago con tripsina, pronasa, DNasa o hialuronidasa. El número de células aisladas depende de las enzimas utilizadas. Así, cuando se trata solo con colagenasa, se pueden obtener de 1,4 a 10⁻¹ condrocitos a partir de 1 g de tejido, mientras que con pronasa, hialuronidasa y colagenasa, se obtienen de 4,3 a10⁻¹. Cuando se trata con colagenasa, el agrecano, las proteínas, la IL-6 y la IL-8 permanecen en el cultivo celular en cantidades significativamente mayores que con el tratamiento secuencial con diferentes enzimas. Existen varias explicaciones para estas diferencias entre los dos cultivos celulares:
- Los receptores celulares son dañados o inhibidos por enzimas. El TGF-beta inhibe la síntesis de ADN y proteoglicanos en condrocitos recién aislados (día 1), mientras que la síntesis de ADN y proteoglicanos en condrocitos cultivados en monocapa (7 días) es estimulada por el TGF-beta. Sin embargo, se requiere un período adecuado para la reexpresión de estos componentes de la membrana antes del inicio del experimento.
- Las proteasas exógenas pueden alterar la interacción célula-matriz mediada por integrinas. La familia de las integrinas promueve la unión de los condrocitos a las moléculas de la matriz extracelular (MEC) (Shakibaei M. et al., 1997). Esta alteración puede afectar la expresión de los genes de la matriz.
- Los restos de los componentes de la matriz pueden regular la función sintética de los condrocitos. Las integrinas son capaces de reconocer los productos de la degradación de la matriz extracelular (ECM), desempeñando así un papel importante en la reparación tisular tras la acción de las enzimas proteolíticas. T. Larsson et al. (1989) informaron que la adición de proteoglicanos intactos o fragmentados al cultivo celular estimula la síntesis de proteínas y proteoglicanos. Sin embargo, un alto nivel de ácido hialurónico causa una disminución significativa en la inclusión de sulfatos en la síntesis de proteoglicanos por condrocitos del embrión de pollo, condrocitos maduros del cerdo y células de condrosarcoma de rata. Además, el ácido hialurónico es un inhibidor de la liberación de proteoglicanos de las células incluso en presencia de IL-1b, TNF-a, FGF, lo que indica la contrarrestación de la primera actividad biológica de los factores de crecimiento y las citocinas. El mecanismo exacto que subyace a la acción del ácido hialurónico sigue sin estar claro; Se sabe que los condrocitos contienen un receptor para el ácido hialurónico asociado a los filamentos de actina del citosol. La unión del ácido hialurónico a su receptor estimula la fosforilación proteica. Por lo tanto, estos datos demuestran la modulación de la función metabólica de los condrocitos por moléculas fragmentadas o nativas de proteínas de la matriz mediante la activación de receptores de la membrana celular.
- La estimulación rápida de la síntesis de proteínas de la matriz por los condrocitos mediante enzimas puede ser una consecuencia de cambios en la forma de los condrocitos y/o en la reorganización del citoesqueleto.
- Algunas citocinas (p. ej., IL-8) y factores de crecimiento (p. ej., IGF-1, TGF-β) se secuestran en la matriz extracelular (MEC). El ejemplo más conocido es la unión del TGF-β con la decorina, lo que resulta en una disminución de su capacidad para inducir el crecimiento celular en células ováricas de hámster chino. El hallazgo de que el contenido de decorina en el cartílago aumenta con la edad sugiere una disminución de la biodisponibilidad del TGF-β con el envejecimiento. Los factores de crecimiento y las citocinas pueden liberarse de los restos de la matriz durante el cultivo y, posteriormente, modular la función de los condrocitos.
Cultivo en monocapa de condrocitos
El fenotipo diferenciado de los condrocitos se caracteriza principalmente por la síntesis de colágeno tipo II y proteoglicanos específicos de tejido, así como por un bajo nivel de actividad mitótica. Existe evidencia de que con el cultivo celular prolongado en una monocapa, así como después de varios pases celulares repetidos, los condrocitos pierden sus contornos esféricos y adquieren una forma alargada similar a la de los fibroblastos. Con dicha metaplasia fibroblástica, la función sintética de las células también se modifica, caracterizándose por una disminución progresiva en la síntesis de colágenos tipos II, IX y XI y un aumento en la síntesis de colágenos tipos I, III e Y. Los proteoglicanos pequeños no agregados se sintetizan debido al agrecano funcional. La síntesis de catepsina B y L es extremadamente baja en las células diferenciadas, pero aumenta en el proceso de pérdida de diferenciación. La colagenasa-1 se expresa en condrocitos diferenciados; Con el cultivo prolongado, su expresión disminuye, mientras que aumenta la producción de inhibidores tisulares de metaloproteasas (TIMP).
Los condrocitos diferenciados reexpresan el colágeno del fenotipo diferenciado al transferirse de una monocapa a un cultivo en suspensión. El proceso de diferenciación probablemente esté relacionado con la forma celular. Esta propiedad es utilizada regularmente por investigadores que estudian injertos defectuosos con condrocitos autólogos. Un pequeño número de células obtenidas de material de biopsia puede expandirse en un cultivo en monocapa y luego reintroducirse en una matriz tridimensional antes del trasplante. La reexpresión de un fenotipo específico por condrocitos desdiferenciados transferidos a un cultivo de agarosa puede ser estimulada por TGF-β, complejo de oseína-hidroxiapatita y ácido ascórbico.
En respuesta a factores de crecimiento y citocinas, los condrocitos se modifican durante el proceso de diferenciación. La respuesta celular a las citocinas y factores de crecimiento difiere entre condrocitos indiferenciados y diferenciados. La IL-1 estimula la proliferación de fibroblastos, mientras que el crecimiento de condrocitos indiferenciados es inhibido por la IL-1. La síntesis de ADN es estimulada por el IGF-1 en condrocitos elongados, pero no en condrocitos aplanados. En condrocitos diferenciados, los efectos estimulantes de la IL-1β y el TNF-α sobre la producción de procolagenasa son más pronunciados que en condrocitos indiferenciados.
Cultivo de condrocitos
El cultivo de condrocitos en suspensión en un medio líquido o en una matriz tridimensional natural o sintética estabiliza el fenotipo condrocítico. Las células conservan su forma esférica y sintetizan proteínas específicas de cada tejido. El cultivo de condrocitos en suspensión se suele recomendar para estudiar la formación de una nueva matriz pericelular. Los cultivos de condrocitos en polímeros absorbentes sintéticos o naturales se utilizan para la implantación de células en defectos cartilaginosos con el fin de estimular la regeneración del tejido cartilaginoso articular. El medio sintético o natural para las células implantadas debe cumplir una serie de requisitos:
- Los implantes deben tener una estructura porosa para la adhesión y el crecimiento celular,
- Ni el polímero en sí ni sus productos de degradación deberían causar inflamación o reacciones tóxicas cuando se implantan in vivo,
- El portador del injerto debe tener la capacidad de unirse al cartílago adyacente o al hueso subcondral,
- La matriz natural o sintética debe tener la capacidad de absorber, su degradación debe ser equilibrada por la regeneración tisular,
- Para facilitar la reparación del cartílago, la estructura química y la arquitectura de los poros de la matriz deben facilitar el mantenimiento del fenotipo celular y la síntesis de proteínas específicas del tejido por parte de los condrocitos colocados en ella.
- Durante la implantación in vivo, es necesario estudiar las propiedades mecánicas de la matriz sintética o natural.
 [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
[ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
Suspensión de condrocitos en fase líquida
La adhesión de células a vasos plásticos en los que se cultivan condrocitos puede prevenirse recubriendo sus paredes con una solución de metilcelulosa, agarosa, hidrogel (poli-2-hidroxietil metacrilato) o una mezcla de colágeno-agarosa. En estas condiciones, los condrocitos forman racimos y sintetizan principalmente agrecano y colágenos específicos de tejido (tipos II, IX, XI). Generalmente se encuentran dos tipos de células. Las células ubicadas en el centro conservan una forma esférica y están rodeadas por una matriz extracelular (ECM) bien desarrollada, lo cual se confirma mediante estudios histoquímicos y ultraestructurales. En la periferia, los condrocitos tienen contornos discoides y están rodeados por una matriz extracelular (ECM) dispersa; se sabe poco sobre las características funcionales de estas células.
Es posible cultivar condrocitos en microtransportadores mantenidos en suspensión; se utilizan como microtransportadores perlas de dextrano (Cytodex), perlas de dextrano recubiertas de colágeno (Cytodex III) y microesferas no porosas de colágeno tipo I (Celagen). En estas condiciones de cultivo, los condrocitos se adhieren a la superficie del microtransportador, conservan su forma esférica y producen un material similar a una matriz. Además, el uso de Celagen promueve la proliferación de condrocitos y la reexpresión del fenotipo normal. Por lo tanto, el cultivo de condrocitos en microesferas de Celagen puede utilizarse para restaurar el fenotipo celular antes del trasplante.
Otro método para cultivar suspensiones de condrocitos en un medio líquido consiste en cultivarlos en forma de bolas densas de células (0,5-1 x 10 b ), obtenidas por centrifugación. Estos condrocitos son capaces de producir una matriz que contiene una gran cantidad de proteoglicanos y colágeno tipo II, pero no de colágeno tipo I, lo cual se confirma mediante métodos histológicos, inmunohistoquímicos y cuantitativos.
Suspensión de condrocitos en ECM natural
Los condrocitos pueden cultivarse en suspensión en una matriz tridimensional (agar blando, agarosa, gel o esponja de colágeno, ácido hialurónico, pegamento de fibrina, perlas de alginato).
Los condrocitos cultivados en agarosa conservan su fenotipo normal y sintetizan colágeno tipo II y agregados de agrecano específicos de tejido. Cuando se cultivan en agarosa, los proteoglicanos sintetizados por la célula se liberan en el medio durante 50 días. A modo de comparación, en un cultivo en monocapa, la fase celular está sobrellenada con glicosaminoglicanos ya en los primeros 5-6 días de cultivo; cuando se cultivan en el medio, después del aumento de la síntesis y liberación de glicosaminoglicanos en los primeros 8-10 días, se produce su disminución dependiente del tiempo. Sin embargo, el comportamiento de los condrocitos cuando se cultivan en agarosa difiere del in vivo. En agarosa, un gran número de agregados de agrecano sintetizados contienen moléculas más pequeñas y menos moléculas que in vivo. TGF-β estimula la síntesis de proteoglicanos en el explante, pero reduce la síntesis de agrecano en agarosa.
El alginato es un polisacárido lineal obtenido de algas pardas. En presencia de cationes divalentes, como los iones Ca₂₂ , este polímero se transforma en un gel. Cada condrocito atrapado en alginato está rodeado por una matriz de polisacáridos con carga negativa, cuyos poros son comparables a los del cartílago hialino. La matriz formada por los condrocitos en las microesferas de alginato consta de dos compartimentos: una fina capa de matriz asociada a las células, correspondiente a la matriz pericelular y territorial del cartílago articular, y una matriz más distante, equivalente a la interterritorial del tejido nativo. Al día 30 de cultivo, el volumen relativo y absoluto ocupado por las células y cada uno de los dos compartimentos en una microesfera de alginato son casi idénticos a los del cartílago nativo. Durante casi 30 días, los condrocitos conservan su forma esférica y producen agrecano, cuyas propiedades hidrodinámicas son similares a las de las moléculas de agrecano en la matriz del cartílago articular, así como a las moléculas de colágeno de los tipos II, IX y XI. Al mismo tiempo, al igual que otros cultivos en suspensión, las células aplanadas están presentes en la superficie de las perlas de alginato, que producen una pequeña cantidad de moléculas de colágeno de tipo I, que se liberan directamente al medio y no se incorporan a la matriz extracelular (ECM). Se observa una proliferación moderada de condrocitos en las perlas de alginato. Después de 8 meses de cultivo en gel de alginato, los condrocitos maduros no pierden actividad metabólica y continúan sintetizando colágeno tipo II y agrecano específicos de tejido.
H. Tanaka et al. (1984) investigaron las propiedades de difusión de diversas moléculas naturales en alginato y descubrieron que las moléculas mayores de 70 kDa no se difundían a través del alginato. Por lo tanto, el cultivo celular en alginato es adecuado para estudiar la regulación de la biosíntesis de la matriz y la organización de la matriz extracelular (ECM). La disponibilidad de células cultivadas en alginato permite estudiar la acción de factores reguladores de péptidos y agentes farmacológicos a nivel transcripcional, postranscripcional y traduccional.
Los condrocitos también se cultivan en una matriz de fibras de colágeno de tipos I y II. S. Nehrer et al. (1997) compararon el funcionamiento de condrocitos caninos en matrices poliméricas porosas de colágeno-proteoglicanos que contenían colágenos de diferentes tipos. Encontraron diferencias importantes en la morfología de la función biosintética de los condrocitos cultivados en matrices de colágeno que contenían colágeno tipos I y II. Las células en una matriz de colágeno tipo II conservaron su forma esférica, mientras que en el colágeno tipo I tenían una morfología similar a la de los fibroblastos. Además, en una matriz de colágeno tipo II, los condrocitos produjeron una mayor cantidad de glicosaminoglicanos. J. van Susante et al. (1995) compararon las propiedades de los condrocitos cultivados en gel de alginato y colágeno (tipo I). Los autores observaron un aumento significativo en el número de células en el gel de colágeno, pero a partir del sexto día de cultivo, las células perdieron su fenotipo característico, transformándose en células similares a fibroblastos. En el gel de alginato, se observó una disminución en el número de células, pero los condrocitos conservaron su fenotipo normal. En el gel de colágeno, el número de proteoglicanos por célula fue significativamente mayor que en el de alginato, pero en el gel se observó una disminución en la síntesis de elementos de la matriz a partir del sexto día de cultivo, mientras que en el de alginato la síntesis continuó aumentando.
La matriz sólida de fibrina tridimensional es una sustancia natural que sustenta a los condrocitos suspendidos en ella en un fenotipo diferenciado. Esta matriz también puede utilizarse como soporte en el trasplante de condrocitos. Las ventajas de la fibrina son la ausencia de citotoxicidad, la capacidad de rellenar espacios y su capacidad adhesiva. Estudios histológicos y bioquímicos, autorradiografía y microscopía electrónica han demostrado que los condrocitos en gel de fibrina conservan su morfología, proliferan y producen matriz incluso después de dos semanas de cultivo. Sin embargo, G. Homminga et al. (1993) informaron que la desintegración de la fibrina comienza después de tres días de cultivo y la desdiferenciación de los condrocitos progresa.
Suspensión de condrocitos en MEC artificial (sintética)
Los implantes de cartílago para cirugía reconstructiva u ortopédica se pueden obtener cultivando condrocitos aislados in vitro en una matriz sintética biocompatible.
Los condrocitos cultivados en ácido poliglicólico proliferan y mantienen una morfología y un fenotipo normales durante ocho semanas. El complejo condrocitos-ácido poliglicólico está compuesto por células, glicosaminoglicanos y colágenos, y posee una cápsula externa de colágeno. Sin embargo, estos implantes contienen dos tipos de moléculas de colágeno: I y II. Los implantes de condrocitos desdiferenciados mediante una serie de pases presentan una mayor cantidad de glicosaminoglicanos y colágenos que los implantes de condrocitos principalmente indiferenciados.
L. Freed et al. (1993b) compararon el comportamiento de cultivos de condrocitos humanos y bovinos en ácido poliglicólico fibroso (FPGA) y ácido poliláctico poroso (PPLA). Después de 6-8 semanas de cultivar condrocitos bovinos en FPGA o PPLA, los autores observaron proliferación celular y regeneración de la matriz de cartílago. En FPGA, los condrocitos tenían forma esférica y se ubicaban en lagunas rodeadas por una matriz de cartílago. Después de 8 semanas de cultivo in vitro, el tejido regenerado contenía hasta un 50% de materia seca (4% de masa celular, 15% de glicosaminoglicanos y 31% de colágenos). En PPLA, las células tenían forma fusiforme y una pequeña cantidad de glicosaminoglicanos y colágeno. En FPGA, el crecimiento celular fue dos veces más intenso que en PPLA. In vivo, los condrocitos cultivados en VPGK y PPLC produjeron tejido histológicamente similar al cartílago en un plazo de 1 a 6 meses. Los implantes contenían glicosaminoglicanos y colágenos tipo I y II.
Se cultivaron condrocitos fetales bovinos en polietileno poroso de alta densidad, tanto hidrofóbico como hidrofílico. Tras 7 días de incubación en ambos sustratos, las células conservaron una forma esférica y contenían principalmente colágeno tipo II. Tras 21 días de cultivo, se observó que la matriz hidrofílica contenía más colágeno tipo II que la matriz hidrofóbica.
El tejido cartilaginoso también puede obtenerse mediante cultivo en monocapa en filtros Millicell-CM. El recubrimiento previo de los filtros con colágeno es necesario para la fijación de las condroitinas. El examen histológico del cultivo demuestra la acumulación de condrocitos en la matriz extracelular (MEC), que contiene proteoglicanos y colágeno tipo II. No se detectó colágeno tipo I en dicho cultivo. Los condrocitos del tejido cartilaginoso obtenido presentan forma esférica, pero en la superficie del tejido están ligeramente aplanados. El grosor del tejido neoformado aumentó con el tiempo y dependió de la densidad inicial de la monocapa celular. En condiciones óptimas de cultivo, el grosor del tejido cartilaginoso alcanzó los 110 μm; la organización de sus células y colágeno en capas superficiales y profundas es similar a la del cartílago articular. La MEC contiene aproximadamente tres veces más colágeno y proteoglicanos. Tras dos semanas de cultivo, se observó acumulación de matriz, lo que permitió extraer el tejido del filtro y utilizarlo para trasplante.
Sims et al. (1996) estudiaron el cultivo de condrocitos en gel de óxido de polietileno, una matriz polimérica encapsulada que permite la transferencia de grandes cantidades de células mediante inyección. Seis semanas después de la inyección en el tejido subcutáneo de ratones atímicos, se formó cartílago nuevo, caracterizado morfológicamente por una opalescencia blanca similar a la del cartílago hialino. Los datos histológicos y bioquímicos indicaron la presencia de condrocitos en proliferación activa que producen matriz extracelular (ECM).
Explantación
La explantación de tejido cartilaginoso se utiliza para estudiar los procesos de anabolismo y catabolismo en él, el mantenimiento de la homeostasis, la resorción y la reparación. Los condrocitos en explantos de cartílago mantienen un fenotipo normal y una composición de la matriz extracelular (ECM) similar a la del cartílago articular in vivo. Tras 5 días de cultivo en presencia de suero, se alcanza un nivel constante de síntesis y procesos de degradación natural. La resorción tisular se puede acelerar en el cultivo principal y en cultivo con la adición de suero utilizando diversos agentes, como IL-1B, TNF-a, lipopolisacáridos bacterianos, derivados del ácido retinoico o radicales activos de oxígeno. Para estudiar la reparación del cartílago, se induce su daño mediante mediadores inflamatorios solubles (H2O2 ,IL -1, TNF-a ) o mediante la ruptura física de la matriz.
El método de cultivo organotípico es un modelo para el estudio in vitro de los efectos de factores externos aislados sobre los condrocitos y la matriz circundante. In vivo, los condrocitos se encuentran dispersamente en la matriz extracelular (MEC) y no entran en contacto entre sí. El cultivo de explantos de cartílago articular preserva esta organización estructural, así como las interacciones específicas entre los condrocitos y el entorno extracelular circundante. Este modelo también se utiliza para estudiar los efectos del estrés mecánico, los agentes farmacológicos, los factores de crecimiento, las citocinas y las hormonas sobre el metabolismo del cartílago.
Otra ventaja de la explantación de tejido cartilaginoso es la ausencia de daño a los condrocitos por la acción de enzimas proteolíticas o factores mecánicos, algo inevitable al aislar células. Los receptores y otras proteínas de membrana y glucoproteínas quedan protegidos de factores dañinos.
[ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ]
Cultivo de condrones
El condrón es una unidad estructural, funcional y metabólica del cartílago articular, compuesta por un condrocito, su matriz pericelular y su cápsula filamentosa compacta, y responsable de la homeostasis de la matriz. Los condrones se extraen mecánicamente del cartílago y se recolectan mediante varias homogeneizaciones sucesivas a baja velocidad. Los condrones aislados de zonas de cartílago con diferentes profundidades se pueden dividir en cuatro categorías: condrón único, condrones pareados, condrones múltiples (tres o más) dispuestos linealmente (columnas de condrones) y cúmulos de condrones.
Los condrones individuales se encuentran generalmente en las capas medias del cartílago intacto, los condrones pareados se encuentran en el límite de las capas media y profunda, los condrones múltiples dispuestos linealmente son típicos de las capas profundas del cartílago intacto. Finalmente, los grupos de condrones consisten en grupos organizados aleatoriamente de condrones individuales y pareados, que conservan un estado agregado después de la homogeneización. Los grupos de condrones son grandes fragmentos de cartílago, que generalmente contienen varios condrones y fibrillas de colágeno dispuestas radialmente, es decir, una organización típica característica de las capas profundas de la matriz. Los condrones se inmovilizan en agarosa transparente, lo que permite estudios de su estructura, composición molecular y actividad metabólica. El sistema condrón-agarosa se considera un micromodelo del cartílago, que difiere del sistema tradicional condrocitos-agarosa en que se conserva el microambiente natural y no hay necesidad de sintetizarlo y ensamblarlo. El cultivo de condrones es un modelo para estudiar las interacciones de las células y la matriz en el cartílago articular en condiciones normales y patológicas.
[ 22 ], [ 23 ], [ 24 ], [ 25 ], [ 26 ], [ 27 ]
Cultivo de condrocitos inmortales
El ADN recombinante o los virus que contienen oncogenes capaces de inmortalizar una célula se utilizan para crear líneas celulares permanentes. Los condrocitos inmortales tienen la capacidad de proliferar indefinidamente manteniendo un fenotipo estable. F. Mallein-Gerin et al. (1995) demostraron que el oncogén SV40T induce la proliferación de condrocitos de ratón, que continúan expresando de forma estable los tipos de colágeno II, IX y XI, así como el agrecano articular y la proteína de unión. Sin embargo, dicha línea celular adquiere la capacidad de sintetizar colágeno tipo I al cultivarla en monocapa o en gel de agarosa.
W. Horton et al. (1988) describieron una línea de células inmortales con un bajo nivel de expresión de ARNm de colágeno tipo II. Estas células se obtuvieron mediante su transformación con un retrovirus de ratón que contenía oncogenes I-myc e y-ra. Este tipo de células representa un modelo único para el estudio de las interacciones de la matriz articular en ausencia de colágeno tipo II, así como la regulación de su síntesis.
El cultivo de condrocitos con genes mutados o delecionados es un modelo conveniente para estudiar su función fisiológica. Este modelo es especialmente adecuado para estudiar el papel de moléculas específicas en la organización de la matriz del cartílago o para investigar los efectos de varios factores reguladores en el metabolismo del cartílago. Los condrocitos con un gen delecionado para el colágeno tipo IX sintetizan fibrillas de colágeno que son más anchas de lo normal, lo que indica que el colágeno tipo IX regula el diámetro de las fibrillas. Como se señaló en el Capítulo 1, recientemente se descubrió una mutación en el gen COLAI que codifica el colágeno tipo II en familias con osteoartritis generalizada primaria. Para estudiar el efecto del colágeno tipo II mutante en la matriz articular, R. Dharmrvaram et al. (1997) transfectaron (infectaron con ácido nucleico extraño) COL 2 AI defectuoso (la arginina en la posición 519 se reemplaza por cisteína) en condrocitos fetales humanos in vitro.
Sistema de cocultivo. En la articulación, el cartílago interactúa con otros tipos de células presentes en la membrana sinovial, el líquido sinovial, los ligamentos y el hueso subcondral. El metabolismo de los condrocitos puede verse afectado por diversos factores solubles sintetizados por estas células. Por lo tanto, en la artritis, el cartílago articular es destruido por enzimas proteolíticas y radicales libres producidos por las células sinoviales. Por ello, se han desarrollado modelos para estudiar las interacciones complejas entre el cartílago y los tejidos circundantes, denominados cocultivos.
S. Lacombe-Gleise et al. (1995) cultivaron condrocitos y osteoblastos de conejo en un sistema de cocultivo (COSTAR), donde las células estaban separadas por una membrana microporosa (0,4 μm), lo que permitía el intercambio entre ambos tipos celulares sin contacto directo. Este estudio demostró la capacidad de los osteoblastos para estimular el crecimiento de los condrocitos mediante mediadores solubles.
AM Malfait y coautores (1994) estudiaron la relación entre monocitos y condrocitos de sangre periférica. Este modelo resulta útil para estudiar los procesos mediados por citocinas en artropatías inflamatorias (artritis reumatoide, espondiloartritis seronegativa, etc.). Los autores del modelo separaron las células con una membrana de unión a proteínas con poros de 0,4 μm de diámetro. El estudio demostró que los monocitos estimulados con lipopolisacárido produjeron IL-1 y TNF-α, lo que inhibió la síntesis de agrecano por los condrocitos y contribuyó a la degradación de los agregados de agrecano ya sintetizados.
K. Tada et al. (1994) crearon un modelo de cocultivo en el que células endoteliales en un gel de colágeno (tipo I) se colocaron en una cámara interna separada de la cámara externa con condrocitos colocados en ella por un filtro con un tamaño de poro de 0,4 μm. En un estado de aislamiento completo de la cámara externa, las células endoteliales humanas formaron tubos en un gel de colágeno en presencia de EGF o TGF-α. Cuando ambos tipos de células se cultivaron simultáneamente, se inhibió la formación de tubos dependiente de TGF-α por las células endoteliales. La inhibición de este proceso por los condrocitos fue parcialmente eliminada por anticuerpos anti-TGF-β. Se puede asumir que el TGF-β producido por los condrocitos inhibe la vascularización del propio cartílago.
S. Groot et al. (1994) cultivaron simultáneamente condrocitos de las zonas hipertrófica y proliferativa del hueso de un feto de ratón de 16 días con fragmentos de tejido cerebral. Tras 4 días de cultivo, se observó la transdiferenciación de condrocitos en osteoblastos y el inicio de la formación de osteoides. Tras 11 días de cultivo, parte del cartílago fue reemplazado por tejido óseo y la matriz ósea se calcificó parcialmente. Algunos neuropéptidos y neurotransmisores producidos por el tejido cerebral afectan al metabolismo de los osteoblastos o tienen receptores para ellos. Entre ellos se encuentran la noradrenalina, el péptido intestinal vasoactivo, el péptido relacionado con el gen de la calcitonina, la sustancia P y la somatostatina. Los fragmentos de tejido cerebral cultivados junto con condrocitos pueden producir algunos de los factores mencionados, capaces de inducir el proceso de transdiferenciación de condrocitos en osteoblastos.
[ 28 ], [ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ]
La influencia de factores externos en el cultivo de condrocitos
La influencia de la tensión del oxígeno en el metabolismo de los condrocitos
En la mayoría de los casos, los cultivos de condrocitos se desarrollan en condiciones de tensión de oxígeno atmosférico. Sin embargo, es bien sabido que los condrocitos in vivo existen en condiciones de hipoxia y que la tensión de oxígeno varía según las condiciones patológicas. Durante el proceso de maduración, se observan cambios significativos en el suministro de sangre a las epífisis. Dado que la vascularización varía en las diferentes zonas de la placa de crecimiento, la tensión de oxígeno en ellas también varía. C. Brighton y R. Heppenstall (1971) demostraron que, en la placa tibial de conejos, la tensión de oxígeno en la zona hipertrófica es menor que en el cartílago circundante. La medición de algunos parámetros metabólicos mostró que los condrocitos pueden responder rápidamente a los cambios locales en la concentración de oxígeno. En primer lugar, a baja tensión de oxígeno, su consumo por los condrocitos disminuye. Con una disminución de la tensión de oxígeno del 21 % al 0,04 %, aumenta la utilización de glucosa, la actividad de las enzimas glucolíticas y la síntesis de ácido láctico. Incluso a baja tensión de oxígeno, la cantidad absoluta de ATP, ADP y AMP se mantiene estable. Estos datos indican que el metabolismo de los condrocitos busca la máxima conservación de energía. Sin embargo, la actividad sintética, y por lo tanto los procesos de reparación, se modifican en condiciones de hipoxia.
La alta tensión de oxígeno también afecta el metabolismo de los condrocitos, provocando una disminución de la síntesis de proteoglicanos y ADN, así como la degradación de la matriz cartilaginosa. Estos efectos suelen ir acompañados de la producción de radicales libres de oxígeno.
La influencia de la concentración de iones y la presión osmótica del entorno en la función de los condrocitos
En el cartílago nativo, la concentración de iones difiere significativamente de la de otros tejidos: el contenido de sodio en el medio extracelular es de 250-350 mmol, y su osmolaridad es de 350-450 mosmol. Cuando se aíslan condrocitos de la matriz extracelular (MEC) y se incuban en un medio estándar (DMEM [Medio Mínimo Esencial de Dulbecco], con una osmolaridad de 250-280,7 mosmol), el entorno celular cambia drásticamente. Además, la concentración de calcio y potasio en el medio estándar es significativamente menor que en el tejido nativo, y la concentración de aniones es significativamente mayor.
La adición de sacarosa al medio aumenta su osmolaridad e induce un aumento intracelular transitorio en la concentración de aniones H + y calcio en el citosol. Dichos cambios intracelulares pueden afectar los procesos de diferenciación de los condrocitos y su actividad metabólica. J. Urban et al. (1993) encontraron que la incorporación de35 8-sulfato y 3 H-prolina por condrocitos aislados incubados en DMEM estándar durante 2-4 h fue solo del 10% de la del tejido nativo. La intensidad de la síntesis alcanzó un máximo a una osmolaridad del medio extracelular de 350-400 mosmol tanto en condrocitos recién aislados como en explantos de tejido cartilaginoso. Además, el volumen de los condrocitos aumentó en un 30-40% después de colocar las células aisladas en DMEM estándar de la osmolaridad especificada. Sin embargo, al cultivar condrocitos en condiciones de osmolaridad no fisiológica durante 12-16 horas, las células se adaptan a las nuevas condiciones, reduciendo la intensidad de la biosíntesis en proporción al cambio de osmolaridad del entorno extracelular.
P. Borgetti et al. (1995) estudiaron el efecto de la osmolaridad del medio extracelular en el crecimiento, la morfología y la biosíntesis de condrocitos porcinos. Los autores demostraron características bioquímicas y morfológicas similares de los condrocitos cultivados en medios con osmolaridad de 0,28 y 0,38 mosmol. Con una osmolaridad del medio de 0,48 mosmol, se observó una disminución de la proliferación celular y la síntesis de proteínas durante las primeras 4-6 horas de cultivo, pero estos parámetros se recuperaron posteriormente y finalmente alcanzaron valores control. Cuando los condrocitos se cultivaron en un medio con una osmolaridad de 0,58 mosmol, las células perdieron la capacidad de mantener la intensidad fisiológica de los procesos proliferativos y, después de 6 días, el número de condrocitos se redujo significativamente. Con una osmolaridad del medio de 0,58 mosmol, se observó una profunda inhibición de la síntesis de proteínas. Además, al cultivarse en medios con una osmolaridad de 0,28-0,38 mOsm, los condrocitos conservan su fenotipo fisiológico; a una osmolaridad mayor (0,48-0,58 mOsm), se producen cambios significativos en la morfología celular, que se manifiestan por la pérdida del fenotipo característico, la transformación de los condrocitos en células similares a fibroblastos y la pérdida de la capacidad celular para ensamblar proteoglicanos de la matriz. Los resultados de este estudio indican la capacidad de los condrocitos para responder a fluctuaciones limitadas en la osmolaridad del entorno extracelular.
Los cambios en la concentración de otros iones también pueden afectar los procesos de biosíntesis en los condrocitos. Así, el grado de incorporación de 35 S (sulfato) aumenta a la mitad con un aumento en la concentración de iones potasio de 5 mmol (la concentración en el medio DM EM estándar) a 10 mmol (la concentración en la MEC in vivo). Las concentraciones de calcio inferiores a 0,5 mmol promovieron la producción de colágeno por condrocitos bovinos maduros, mientras que una concentración de 1-2 mmol (correspondiente a la concentración en el medio DM EM estándar) causó una disminución significativa en la síntesis de colágeno. Se observó un aumento moderado en la biosíntesis a altos niveles de calcio (2-10 mmol). Varios cationes participan en la unión de los condrocitos a las proteínas de la MEC. Así, los iones de magnesio y manganeso proporcionan unión a la fibronectina y al colágeno tipo II, mientras que los iones de calcio no participan en la unión de los condrocitos a las proteínas. Así, los resultados de los estudios descritos indican la influencia de los cambios en los iones extracelulares de potasio, sodio, calcio y la osmolaridad del medio sobre la función biosintética de los condrocitos incubados en medios estándar.
Efecto del estrés mecánico sobre el metabolismo de los condrocitos
La inmovilización articular provoca atrofia reversible del cartílago, lo que indica la necesidad de estímulos mecánicos para los procesos metabólicos normales en la matriz extracelular (ECM). En la mayoría de los casos, los modelos de cultivo celular utilizados se encuentran bajo presión atmosférica normal. M. Wright et al. (1996) demostraron que el entorno mecánico afecta el metabolismo de los condrocitos; la respuesta celular depende de la intensidad y frecuencia de la carga compresiva. Experimentos in vitro con carga en explantos de cartílago articular intacto demostraron una disminución en la síntesis de proteínas y proteoglicanos bajo la acción de la carga estática, mientras que la carga dinámica estimula estos procesos. Los mecanismos exactos del efecto de la carga mecánica sobre el cartílago son complejos y probablemente estén asociados con la deformación celular, la presión hidrostática, la presión osmótica, el potencial eléctrico y los receptores celulares superficiales para las moléculas de la matriz. Para estudiar el efecto de cada uno de estos parámetros, es necesario crear un sistema en el que cada parámetro pueda variarse de forma independiente. Por ejemplo, el cultivo de explantos no es adecuado para estudiar la deformación celular, pero sí puede utilizarse para estudiar el efecto general de la presión sobre la actividad metabólica de los condrocitos. La compresión del cartílago provoca la deformación celular y se acompaña de la aparición de un gradiente de presión hidrostática, potencial eléctrico, flujo de fluidos y cambios en parámetros fisicoquímicos como el contenido de agua en la matriz, la densidad de carga eléctrica y el nivel de presión osmótica. La deformación celular puede estudiarse utilizando condrocitos aislados sumergidos en gel de agarosa o colágeno.
Se han desarrollado varios sistemas para estudiar el efecto de la estimulación mecánica en el cultivo de condrocitos. Algunos investigadores utilizan sistemas en los que se aplica presión al cultivo celular a través de la fase gaseosa. Así, JP Veldhuijzen et al. (1979), utilizando una presión de 13 kPa por encima de la atmosférica con una frecuencia baja (0,3 Hz) durante 15 min, observaron un aumento en la síntesis de AMPc y proteoglicanos y una disminución en la síntesis de ADN. R. Smith et al. (1996) demostraron que la exposición intermitente de un cultivo de condrocitos bovinos primarios a presión hidrostática (10 MPa) con una frecuencia de 1 Hz durante 4 h provocó un aumento en la síntesis de agrecano y colágeno tipo II, mientras que la presión constante no afectó estos procesos. Utilizando un sistema similar, Wright et al. (1996) informaron que la presión cíclica en el cultivo celular está asociada con la hiperpolarización de la membrana celular del condrocitos y la activación de los canales de potasio dependientes de Ca 2+. Por lo tanto, los efectos de la presión cíclica están mediados por canales iónicos activados por estiramiento en la membrana del condrocitos. La respuesta de los condrocitos a la presión hidrostática depende de las condiciones del cultivo celular y de la frecuencia de la carga aplicada. Así, la presión hidrostática cíclica (5 MPa) disminuye la incorporación de sulfato a la monocapa de condrocitos a una frecuencia de 0,05, 0,25 y 0,5 Hz, mientras que a una frecuencia superior a 0,5 Hz, aumenta la incorporación de sulfato al explante de cartílago.
M. Bushmann et al. (1992) informaron que los condrocitos en geles de agarosa alteran su biosíntesis en respuesta a cargas mecánicas estáticas y dinámicas, de la misma manera que el órgano intacto cultivado. Los autores observaron que la carga mecánica genera un estímulo hiperosmótico con la consiguiente disminución del pH en los condrocitos.
El efecto del estiramiento mecánico puede estudiarse en un cultivo celular sumergido en un gel. La fuerza de estiramiento puede crearse utilizando un vacío controlado por computadora. Cuando el sistema está bajo cierto grado de vacío, el fondo de la placa de Petri con el cultivo celular se extiende por una cantidad conocida, la deformación es máxima en los bordes del fondo de la placa y mínima en el centro. El estiramiento también se transmite a los condrocitos cultivados en la placa de Petri. Usando este método, K. Holm-vall et al. (1995) demostraron que en células de condrosarcoma cultivadas en un gel de colágeno (tipo II), la expresión de ARNm de una integrina 2 aumenta. Una integrina2 β es capaz de unirse al colágeno tipo II. Se considera un mecanorreceptor, ya que interactúa con proteínas de unión a actina, conectando así la matriz extracelular (ECM) y el citoesqueleto.
El efecto del pH en el metabolismo de los condrocitos
El pH del líquido intersticial de la matriz extracelular (ECM) del cartílago es más ácido que en otros tejidos. A. Maroudas (1980) determinó el pH de la matriz del cartílago articular en 6,9. B. Diamant et al. (1966) encontraron un pH de 5,5 en condiciones patológicas. Se sabe que los condrocitos viven con baja PO2, lo que indica el importante papel de la glucólisis (95 % del metabolismo total de la glucosa) en el metabolismo de estas células; la glucólisis se acompaña de la producción de una gran cantidad de ácido láctico.
Además de la acidificación del entorno por los productos de la glucólisis, los propios componentes de la matriz son de gran importancia. La gran cantidad de carga negativa fija en los proteoglicanos modifica la composición iónica extracelular: se observa una alta concentración de cationes libres (p. ej., H +, Na +, K + ) y una baja concentración de aniones (p. ej., O2, HCO3). Además, bajo la influencia de la carga mecánica, se expulsa agua de la MEC, lo que conduce a un aumento en la concentración de cargas negativas fijas y a la atracción de más cationes hacia la matriz. Esto se acompaña de una disminución del pH del entorno extracelular, que afecta al pH intracelular, modificando así el metabolismo de los condrocitos. R. Wilkin y A. Hall (1995) estudiaron el efecto del pH del entorno extracelular e intracelular en la biosíntesis de la matriz por condrocitos bovinos aislados. Observaron una modificación dual de la síntesis de la matriz con una disminución del pH. Una ligera disminución del pH (7,4
[ 34 ], [ 35 ], [ 36 ], [ 37 ], [ 38 ], [ 39 ], [ 40 ]
Influencia de la composición del medio de cultivo en el metabolismo de los condrocitos
El medio de cultivo de condrocitos debe ser adecuado para las condiciones experimentales. En los últimos años, se ha utilizado suero bovino para optimizar las condiciones de cultivo. Sin embargo, al utilizar suero, deben tenerse en cuenta varios puntos importantes:
- crecimiento externo de células desde la periferia del tejido en cultivos de órganos,
- variabilidad en la composición de sueros de diferentes series,
- la presencia de componentes desconocidos en ellos,
- Mayor riesgo de interferencias y artefactos al estudiar la influencia de diversos factores biológicos en la actividad metabólica de las células.
Un ejemplo de esto último es un estudio sobre el efecto del EGF en los condrocitos del cartílago de ratas. El EGF estimuló la incorporación de 3H -timidina y un aumento del contenido de ADN en el cultivo. Este efecto fue más pronunciado a bajas concentraciones séricas (<1%), pero a altas concentraciones (>7,5%) desapareció.
Es bien conocido que los niveles de síntesis y degradación en DMEM suplementado con suero bovino aumentan significativamente en comparación con las condiciones in vivo. Las diferencias entre el metabolismo in vivo e in vitro pueden deberse a diferencias entre el líquido sinovial y el medio en el que se cultivan las células. Lee et al. (1997) cultivaron condrocitos bovinos jóvenes en agarosa utilizando un medio nutritivo que contenía DMEM suplementado con 20% de suero bovino y una gran cantidad de líquido sinovial alogénico normal. La presencia de líquido sinovial en el medio indujo un aumento en la cantidad de proteoglicanos, hasta el 80% de la cantidad total de líquido sinovial. Estos resultados indican que el líquido sinovial en cultivo induce un nivel de metabolismo similar al in vivo, con un alto nivel de síntesis de glicosaminoglicanos y un bajo nivel de división celular.
G. Verbruggen et al. (1995) demostraron que la síntesis de ³²S -arrpeKaHa por condrocitos humanos cultivados en agarosa en DMEM sin suero fue del 20-30 % del nivel de síntesis observado en DMEM suplementado con un 10 % de suero bovino. Los autores determinaron hasta qué punto IGF-1, IGF-2, TGF-R o insulina restauraron la producción de agrecano en medio sin suero. Concluyeron que 100 ng/ml de insulina, IGF-1 o IGF-2 restauraron parcialmente la síntesis de agrecano al 39-53 % del nivel de control. No se observó sinergia ni acumulación con una combinación de los factores mencionados. Al mismo tiempo, 10 ng/ml de TGF-R en presencia de 100 ng/ml de insulina estimularon la síntesis de agrecano al 90 % o más del nivel de referencia. Finalmente, la transferrina sérica humana, sola o en combinación con insulina, no afectó la síntesis de agrecano. Cuando el suero bovino se reemplazó con albúmina sérica bovina, el contenido de agregados de agrecano disminuyó significativamente. El enriquecimiento del medio de cultivo con insulina, IGF o TGF-R restauró parcialmente la capacidad de las células para producir agregados de agrecano. Además, el IGF-1 y la insulina pueden mantener la homeostasis en cultivos celulares. Después de 40 días de cultivo en un medio enriquecido con 10-20 ng/ml de IGF-1, la síntesis de proteoglicanos se mantuvo al mismo nivel o incluso más alto en comparación con el medio que contenía suero bovino al 20%. Los procesos catabólicos procedieron más lentamente en el medio enriquecido con IGF-1 que en el medio enriquecido con solución de albúmina al 0,1%, pero algo más rápido en el medio enriquecido con suero al 20%. En cultivos de larga vida, 20 ng/ml de IGF-1 mantienen un estado estable de las células.
D. Lee et al. (1993) compararon el efecto de la composición del medio de cultivo (DMEM, DMEM + 20% suero bovino, DMEM + 20 ng/ml IGF-1) sobre la síntesis de ADN en un cultivo de explante de tejido de cartílago, un cultivo en monocapa y en suspensión de agarosa. Al cultivar en agarosa en presencia de suero, los autores observaron una tendencia de los condrocitos a agruparse en grandes grupos. Las células cultivadas sin suero o con IGF-1 conservaron una forma redonda en agarosa, se recolectaron en pequeños grupos, pero no formaron agregados grandes. En una monocapa, la síntesis de ADN fue significativamente mayor en el medio que contenía suero que en el medio enriquecido con IGF-1; la síntesis de ADN en este último fue significativamente mayor que en el medio no enriquecido. No se encontraron diferencias en la síntesis de ADN cuando los condrocitos se cultivaron en suspensión de agarosa en un medio no enriquecido y en un medio con IGF-1. Al mismo tiempo, el cultivo de suspensiones de condrocitos en agarosa en un medio enriquecido con suero estuvo acompañado de una mayor incorporación del radionucleótido 3H -timidina en comparación con otros medios.
La vitamina C es necesaria para la activación de las enzimas implicadas en la formación de una estructura helicoidal estable en las fibrillas de colágeno. Los condrocitos deficientes en ácido ascórbico sintetizan precursores de colágeno no helicoidales subhidroxilados, que se secretan lentamente. La administración de ácido ascórbico (50 μg/ml) provoca la hidroxilación de los colágenos de tipo II y IX y su secreción en cantidades normales. La adición de vitamina C no afectó la síntesis de proteoglicanos. Por lo tanto, la secreción de colágeno se regula independientemente de la secreción de proteoglicanos.