Médico experto del artículo.

Nuevos artículos
Chlamydiae
Último revisado: 06.07.2025

Todo el contenido de iLive se revisa médicamente o se verifica para asegurar la mayor precisión posible.
Tenemos pautas de abastecimiento estrictas y solo estamos vinculados a sitios de medios acreditados, instituciones de investigación académica y, siempre que sea posible, estudios con revisión médica. Tenga en cuenta que los números entre paréntesis ([1], [2], etc.) son enlaces a estos estudios en los que se puede hacer clic.
Si considera que alguno de nuestros contenidos es incorrecto, está desactualizado o es cuestionable, selecciónelo y presione Ctrl + Intro.
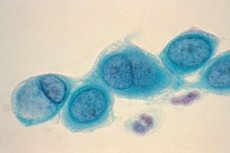
Las clamidias son pequeñas bacterias parasitarias cocoides gramnegativas pertenecientes al orden Chlamydiales, familia Chlamydiaceae. Actualmente, esta familia incluye dos géneros que difieren en estructura antigénica, inclusiones intracelulares y sensibilidad a las sulfonamidas: Chlamydia ( Chlamydia trachomatis ) y Chlamydophila (Chlamydia pneumoniae, Chlamydia psittaci ).
El nombre "clamidia" (del griego chtamys - manto) refleja la presencia de una membrana alrededor de las partículas microbianas.
Todos los tipos de clamidia comparten características morfológicas, un antígeno de grupo común y un ciclo reproductivo específico. Se consideran bacterias gramnegativas que han perdido la capacidad de sintetizar ATP. Por lo tanto, son parásitos intracelulares obligados a generar energía.
Chlamydia trachomatis y Chlamydia pneumoniae se clasifican como microorganismos definitivamente patógenos para los humanos y son los agentes causantes de la clamidia antropogénica. Según el tipo de patógeno y la vía de entrada (tracto respiratorio, aparato genitourinario), se distinguen la clamidia respiratoria y la urogenital.
Se han descrito más de 20 formas nosológicas causadas por Chlamydia trachomatis, entre ellas tracoma, conjuntivitis, linfogranulomatosis inguinal, síndrome de Reiter y clamidia urogenital. Según estimaciones de la OMS, las infecciones por Chlamydia trachomatis ocupan el segundo lugar entre las enfermedades de transmisión sexual, después de las infecciones por tricomonas. Cada año se registran alrededor de 50 millones de casos en todo el mundo.
La neumonía por Chlamydophila causa neumonía grave, una enfermedad de las vías respiratorias superiores. Se ha sugerido que esta bacteria está implicada en el desarrollo de la aterosclerosis y el asma bronquial.
Chlamydophila psittaci es la causa de la ornitosis (psitacosis), una enfermedad zoonótica.
Propiedades morfológicas y tintóreas de la clamidia
Las clamidias son pequeñas bacterias gramnegativas de forma esférica u ovoide. Carecen de flagelos y cápsulas. El principal método para detectarlas es la tinción de Romanovsky-Giemsa. El color de la tinción depende de la etapa del ciclo biológico: los anillos elementales se tiñen de púrpura sobre un fondo azul del citoplasma celular, y los cuerpos reticulares de azul.
La estructura de la pared celular se asemeja a la de las bacterias gramnegativas, aunque presenta diferencias. Carece del típico péptido glicano: el ácido N-acetilmurámico está completamente ausente en su composición. La pared celular incluye una membrana externa que contiene LPS y proteínas. A pesar de la ausencia de péptido glicano, la pared celular de la clamidia es rígida. El citoplasma celular está delimitado por una membrana citoplasmática interna.
El análisis de la membrana externa (MO) de la clamidia mostró que contiene LPS, la proteína principal de la membrana externa (MOMP), y las proteínas Ompl y Omp3, ricas en cisteína, asociadas con la superficie interna de la MO. El LPS y la MOMP de Chlamydia psittaci y Chlamydia trachomatis, a diferencia de la MOMP de Chlamydia pneumoniae, se localizan en la superficie externa de la célula. Las proteínas Omp de Chlamydia psittaci y Chlamydia pneumoniae, con un peso molecular de 90-100 kDa, también se encuentran aquí.
Las clamidias son polimórficas, lo que se relaciona con las peculiaridades de su reproducción. Su singular ciclo de desarrollo (bifásico) se caracteriza por la alternancia de dos formas de existencia: la infecciosa (cuerpos elementales - CE) y la vegetativa (cuerpos reticulares o iniciales - RT).
Los microorganismos contienen ARN y ADN. En la RT, el ARN es cuatro veces mayor que el ADN. En estos, el contenido es equivalente.
Los cuerpos reticulares pueden ser ovalados, con forma de medialuna, en forma de bastoncillos bipolares y cocobacilos, con un tamaño de 300 a 1000 nm. Los cuerpos reticulares no presentan propiedades infecciosas y, al dividirse, facilitan la reproducción de la clamidia.
Los cuerpos elementales, de forma ovalada y con un tamaño de 250-500 nm, poseen propiedades infecciosas y pueden penetrar en una célula sensible, donde se desarrolla el ciclo celular. Poseen una membrana externa densa que los hace resistentes en el medio extracelular.
Cultivo de clamidia
Las clamidias, al ser parásitos obligados, no se reproducen en medios nutritivos artificiales; solo pueden cultivarse en células vivas. Son parásitos energéticos, ya que no pueden acumular energía de forma independiente ni utilizar el ATP de la célula huésped. Las clamidias se cultivan en cultivos de células HeLa y McCoy, en sacos vitelinos de embriones de pollo y en el cuerpo de animales sensibles a una temperatura de 35 °C.
 [ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ]
[ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ]
Estructura antigénica de la clamidia
La clamidia tiene tres tipos de antígenos: antígeno específico (común a todos los tipos de clamidia) - LPS; antígeno infoespecífico (diferente para todos los tipos de clamidia) - proteína de naturaleza, ubicada en la membrana externa; específico de tipo (diferente para los serovares de Chlamydia trachomatis) - LPS, que se multiplica en la pared celular del microorganismo; antígeno variante-específico de naturaleza proteica.
Los serovares A, B y C se denominan oculares, ya que causan tracoma. Los serovares D, E, K, O, H, I, J y K (genial) son los agentes causales de la clamidia urogenital y sus complicaciones. El serovare L es el agente causal de la linfogranulomatosis venérea. El agente causal de la clamidia respiratoria, Chlamydia pneumoniae, tiene 4 serovares: TWAR, AR, RF y CWL. Chlamydia psittaci tiene 13 serovares.
[ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
Tropismo celular de la clamidia
Chlamydia trachomatis tiene tropismo por la mucosa del epitelio del tracto urogenital y puede permanecer localmente en ella o extenderse por toda la superficie del tejido. El agente causal del linfogranuloma venéreo tiene tropismo por el tejido linfoide.
Chlamydia pneumoniae se replica en macrófagos alveolares, monocitos y células endoteliales vasculares; también es posible la diseminación sistémica de la infección.
Chlamydia psittaci causa infección en una variedad de tipos de células, incluidos los fagocitos mononucleares.
Ciclo de vida de la clamidia
El ciclo de desarrollo de la clamidia dura 40-72 horas e incluye dos formas de existencia que difieren en propiedades morfológicas y biológicas.
En la primera etapa del proceso infeccioso, la adsorción de cuerpos elementales de clamidia en la membrana plasmática de la célula huésped sensible se produce mediante fuerzas electrostáticas. La introducción de clamidia en la célula se produce por endocitosis. Las secciones de la membrana plasmática con cuerpos elementales adsorbidos se invaginan en el citoplasma, formando vacuolas fagocíticas. Esta etapa dura de 7 a 10 horas.
Luego, en un plazo de 6 a 8 horas, los cuerpos elementales infecciosos se reorganizan en formas intracelulares vegetativas, metabólicamente activas y no infecciosas (RT), que se dividen repetidamente. Estas formas intracelulares, que son microcolonias, se denominan inclusiones de clamidia. Entre 18 y 24 horas después de su desarrollo, se localizan en una vesícula citoplasmática formada a partir de la membrana de la célula huésped. La inclusión puede contener de 100 a 500 cuerpos reticulares de clamidia.
En la siguiente etapa, durante 36-42 horas, se produce la maduración (formación de cuerpos intermedios) y la transformación de los cuerpos reticulares por división en cuerpos elementales. La célula infectada se destruye y los cuerpos elementales la abandonan. Al ser extracelulares, los cuerpos elementales penetran en nuevas células huésped después de 40-72 horas, comenzando así un nuevo ciclo de desarrollo de la clamidia.
Además de dicho ciclo reproductivo, en condiciones desfavorables se producen otros mecanismos de interacción entre la clamidia y la célula huésped. Estos incluyen la destrucción de la clamidia en los fagosomas, la transformación tipo L y la persistencia.
Las formas transformadas y persistentes de clamidia son capaces de volver a las formas originales (reticulares) con posterior transformación en cuerpos elementales.
Fuera de las células huésped, las funciones metabólicas se reducen al mínimo.
Factores de patogenicidad de la clamidia
Las propiedades adhesivas de la clamidia se deben a las proteínas de la membrana externa de las células, que también poseen propiedades antifagocíticas. Además, las células microbianas poseen endotoxinas y producen exotoxinas. Las endotoxinas están representadas por LPS, similares en muchos aspectos al LPS de las bacterias gramnegativas. La termolabilidad de la sustancia se debe a las exotoxinas, presentes en todos los ratones y que causan la muerte tras su administración intravenosa.
Se ha descubierto que la clamidia tiene un sistema secretor tipo III, a través del cual las proteínas clamidiales se inyectan en el citoplasma de la célula huésped como parte integral del proceso infeccioso.
La proteína de choque térmico (HSP) tiene la capacidad de inducir reacciones autoinmunes.
[ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ], [ 34 ]
Ecología y resistencia de la clamidia
Las clamidias son microorganismos muy comunes. Se han encontrado en más de 200 especies de animales, peces, anfibios, moluscos y artrópodos. También se han encontrado microorganismos con morfología similar en plantas superiores. Los principales hospedadores de las clamidias son los humanos, las aves y los mamíferos.
El agente causal de la clamidia es inestable en el ambiente externo, muy sensible a las altas temperaturas y muere rápidamente al secarse. Su inactivación a 50 °C ocurre después de 30 minutos, a 90 °C, después de 1 minuto. A temperatura ambiente (18-20 °C), la actividad infecciosa del patógeno disminuye después de 5-7 días. A 37 °C, se observa una disminución de la virulencia del 80% después de 6 horas en un termostato. Las bajas temperaturas (-20 °C) contribuyen a la preservación a largo plazo de las propiedades infecciosas del patógeno. La clamidia muere rápidamente bajo la influencia de la radiación UV, por contacto con éter etílico y etanol al 70%, bajo la influencia de lisol al 2% durante 10 minutos y cloramina al 2%.