Médico experto del artículo.

Nuevos artículos
glándula pituitaria
Último revisado: 04.07.2025

Todo el contenido de iLive se revisa médicamente o se verifica para asegurar la mayor precisión posible.
Tenemos pautas de abastecimiento estrictas y solo estamos vinculados a sitios de medios acreditados, instituciones de investigación académica y, siempre que sea posible, estudios con revisión médica. Tenga en cuenta que los números entre paréntesis ([1], [2], etc.) son enlaces a estos estudios en los que se puede hacer clic.
Si considera que alguno de nuestros contenidos es incorrecto, está desactualizado o es cuestionable, selecciónelo y presione Ctrl + Intro.
La glándula pituitaria (hipófisis, s. glandula pituitaria) se encuentra en la fosa hipofisaria de la silla turca del hueso esfenoides y está separada de la cavidad craneal por un proceso de la duramadre cerebral, formando el diafragma de la silla turca. A través de la abertura en este diafragma, la glándula pituitaria se conecta al infundíbulo del hipotálamo del diencéfalo. El tamaño transversal de la glándula pituitaria es de 10-17 mm, el anteroposterior es de 5-15 mm, el vertical es de 5-10 mm. La masa de la glándula pituitaria en hombres es de aproximadamente 0,5 g, en mujeres - 0,6 g. La glándula pituitaria está cubierta externamente por una cápsula.
De acuerdo con el desarrollo de la glándula pituitaria a partir de dos rudimentos diferentes, se distinguen dos lóbulos en el órgano: el anterior y el posterior. La adenohipófisis, o lóbulo anterior (adenohipófisis, s.lobus anterior), es más grande y constituye el 70-80% de la masa total de la glándula pituitaria. Es más densa que el lóbulo posterior. En el lóbulo anterior, se distingue una parte distal (pars distalis), que ocupa la parte anterior de la fosa pituitaria, una parte intermedia (pars intermedia), ubicada en el límite con el lóbulo posterior, y una parte tuberosa (pars tuberalis), que asciende y se conecta con el infundíbulo del hipotálamo. Debido a la abundancia de vasos sanguíneos, el lóbulo anterior tiene un color amarillo pálido con un tinte rojizo. El parénquima de la glándula pituitaria anterior está representado por varios tipos de células glandulares, entre cuyas hebras se ubican capilares sanguíneos sinusoidales. El 50 % de las células de la adenohipófisis son adenocitos cromófobos, que presentan gránulos finos en su citoplasma que se tiñen bien con sales de cromo. Estos son adenocitos acidófilos (40 % de todas las células de la adenohipófisis) y adenocitos basófilos (10 %). Los adenocitos basófilos incluyen endocrinocitos gonadotrópicos, corticotrópicos y tirotrópicos. Los adenocitos cromófobos son pequeños, tienen un núcleo grande y una pequeña cantidad de citoplasma. Estas células se consideran precursoras de los adenocitos cromófobos. El 50 % restante de las células de la adenohipófisis son adenocitos cromófobos.

La neurohipófisis, o lóbulo posterior (neurohipófisis, s.lobus posterior), está compuesta por el lóbulo neural (lobus nervosus), ubicado en la parte posterior de la fosa hipofisaria, y el infundíbulo, ubicado detrás de la porción tuberosa de la adenohipófisis. El lóbulo posterior de la hipófisis está formado por células neurogliales (células hipofisarias), fibras nerviosas que van desde los núcleos neurosecretores del hipotálamo hasta la neurohipófisis, y corpúsculos neurosecretores.
La hipófisis, a través de fibras nerviosas (vías) y vasos sanguíneos, está conectada funcionalmente con el hipotálamo del diencéfalo, que regula su actividad. La hipófisis y el hipotálamo, junto con sus conexiones neuroendocrinas, vasculares y nerviosas, suelen considerarse el sistema hipotálamo-hipofisario.
Las hormonas de la hipófisis anterior y posterior afectan a muchas funciones del cuerpo, principalmente a través de otras glándulas endocrinas. En la hipófisis anterior, los adenocitos acidófilos (células alfa) producen la hormona somatotrópica (hormona del crecimiento), que participa en la regulación de los procesos de crecimiento y desarrollo en organismos jóvenes. Los endocrinocitos corticotrópicos secretan la hormona adrenocorticotrópica (ACTH), que estimula la secreción de hormonas esteroides por las glándulas suprarrenales. Los endocrinocitos tirotrópicos secretan la hormona tirotrópica (TSH), que afecta al desarrollo de la glándula tiroides y activa la producción de sus hormonas. Las hormonas gonadotrópicas: folículo estimulante (FSH), luteinizante (LH) y prolactina, afectan la maduración sexual del cuerpo, regulan y estimulan el desarrollo de folículos en el ovario, la ovulación, el crecimiento de las glándulas mamarias y la producción de leche en las mujeres, y el proceso de espermatogénesis en los hombres. Estas hormonas son producidas por los adenocitos basófilos (células beta). Aquí también se secretan factores lipotrópicos de la hipófisis, que influyen en la movilización y utilización de las grasas en el organismo. En la parte intermedia del lóbulo anterior se produce la hormona estimulante de los melanocitos, que controla la formación de pigmentos (melaninas) en el organismo.
Las células neurosecretoras de los núcleos supraóptico y paraventricular del hipotálamo producen vasopresina y oxitocina. Estas hormonas se transportan a las células de la hipófisis posterior a través de los axones que conforman el tracto hipotálamo-hipofisario. Desde la hipófisis posterior, estas sustancias pasan a la sangre. La hormona vasopresina tiene un efecto vasoconstrictor y antidiurético, por lo que también se denomina hormona antidiurética (ADH). La oxitocina estimula la contractilidad de los músculos uterinos, aumenta la secreción de leche por la glándula mamaria lactante, inhibe el desarrollo y la función del cuerpo lúteo y modifica el tono de la musculatura lisa (no estriada) del tracto gastrointestinal.
Desarrollo de la glándula pituitaria
El lóbulo anterior de la hipófisis se desarrolla a partir del epitelio de la pared dorsal del arco oral en forma de una excrecencia anular (bolsa de Rathke). Esta protuberancia ectodérmica crece hacia la base del futuro tercer ventrículo. Hacia ella, desde la superficie inferior de la segunda vesícula cerebral (la futura base del tercer ventrículo), crece una prolongación, a partir de la cual se desarrollan el tubérculo gris del infundíbulo y el lóbulo posterior de la hipófisis.
Vasos y nervios de la glándula pituitaria
Desde las arterias carótidas internas y los vasos del círculo arterial del cerebro, las arterias pituitarias superior e inferior se dirigen a la glándula pituitaria. Las arterias pituitarias superiores van al núcleo gris y al infundíbulo del hipotálamo, se anastomosan entre sí aquí y forman capilares que penetran en el tejido cerebral: la red hemocapilar primaria. A partir de los bucles largos y cortos de esta red, se forman las venas porta, que se dirigen al lóbulo anterior de la glándula pituitaria. En el parénquima del lóbulo anterior de la glándula pituitaria, estas venas se dividen en amplios capilares sinusoidales, formando la red hemocapilar secundaria. El lóbulo posterior de la glándula pituitaria se irriga principalmente por la arteria pituitaria inferior. Hay largas anastomosis arteriales entre las arterias pituitarias superior e inferior. La salida de sangre venosa de la red hemocapilar secundaria se realiza a través de un sistema de venas que desembocan en los senos cavernosos e intercavernosos de la duramadre del cerebro.
Las fibras simpáticas que entran en el órgano junto con las arterias participan en la inervación de la hipófisis. Las fibras nerviosas simpáticas posganglionares parten del plexo de la arteria carótida interna. Además, numerosas terminaciones de las prolongaciones de las células neurosecretoras ubicadas en los núcleos del hipotálamo se encuentran en el lóbulo posterior de la hipófisis.
Características de la glándula pituitaria relacionadas con la edad
El peso promedio de la hipófisis en los recién nacidos es de 0,12 g. El peso de este órgano se duplica a los 10 años y se triplica a los 15. A los 20 años, el peso de la hipófisis alcanza su máximo (530-560 mg) y se mantiene prácticamente sin cambios en las etapas posteriores. Después de los 60 años, se observa una ligera disminución del peso de esta glándula endocrina.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hormonas pituitarias
La unidad de la regulación nerviosa y hormonal en el cuerpo está asegurada por la estrecha conexión anatómica y funcional entre la hipófisis y el hipotálamo. Este complejo determina el estado y el funcionamiento de todo el sistema endocrino.
La principal glándula endocrina que produce varias hormonas peptídicas que regulan directamente la función de las glándulas periféricas es la hipófisis. Es una formación de color gris rojizo con forma de frijol cubierta por una cápsula fibrosa que pesa entre 0,5 y 0,6 g. Varía ligeramente según el sexo y la edad de la persona. La división generalmente aceptada de la hipófisis en dos lóbulos que difieren en desarrollo, estructura y función se mantiene: el distal anterior (adenohipófisis) y el posterior (neurohipófisis). El primero constituye aproximadamente el 70% de la masa total de la glándula y se divide convencionalmente en las partes distal, infundibular e intermedia, el segundo en la parte posterior, o lóbulo, y el tallo hipofisario. La glándula está ubicada en la fosa hipofisaria de la silla turca del hueso esfenoides y está conectada al cerebro a través del tallo. La parte superior del lóbulo anterior está cubierta por el quiasma óptico y los tractos ópticos. El suministro de sangre a la glándula pituitaria es muy abundante y se lleva a cabo por ramas de la arteria carótida interna (las arterias pituitarias superior e inferior), así como por ramas del círculo arterial del cerebro. Las arterias pituitarias superiores participan en el suministro de sangre a la adenohipófisis, y las inferiores, a la neurohipófisis, contactando con las terminaciones neurosecretoras de los axones de los núcleos de células grandes del hipotálamo. Las primeras ingresan a la eminencia media del hipotálamo, donde se dispersan en una red capilar (el plexo capilar primario). Estos capilares (con los que contactan las terminales de los axones de las pequeñas células neurosecretoras del hipotálamo mediobasal) se agrupan en venas porta que descienden a lo largo del tallo pituitario hacia el parénquima de la adenohipófisis, donde nuevamente se dividen en una red de capilares sinusoidales (el plexo capilar secundario). Así, la sangre, habiendo pasado previamente por la eminencia media del hipotálamo, donde se enriquece con hormonas adenohipofisótropas hipotalámicas (hormonas liberadoras), entra en la adenohipófisis.
La salida de sangre saturada de hormonas adenohipofisarias desde los numerosos capilares del plexo secundario se realiza a través del sistema venoso, que a su vez desemboca en los senos venosos de la duramadre y, posteriormente, en el torrente sanguíneo general. Así pues, el sistema porta de la glándula pituitaria, con un flujo sanguíneo descendente desde el hipotálamo, es un componente morfofuncional del complejo mecanismo de control neurohumoral de las funciones trópicas de la adenohipófisis.
La hipófisis está inervada por fibras simpáticas que siguen las arterias hipofisarias. Estas se originan en fibras posganglionares que atraviesan el plexo carotídeo interno, conectado a los ganglios cervicales superiores. La adenohipófisis no recibe inervación directa del hipotálamo. El lóbulo posterior recibe fibras nerviosas de los núcleos neurosecretores del hipotálamo.
La adenohipófisis es una formación muy compleja en su arquitectura histológica. Presenta dos tipos de células glandulares: cromófobas y cromófilas. Estas últimas se dividen a su vez en acidófilas y basófilas (se ofrece una descripción histológica detallada de la glándula pituitaria en la sección correspondiente del manual). Sin embargo, cabe destacar que las hormonas producidas por las células glandulares que conforman el parénquima de la adenohipófisis, debido a la diversidad de este último, difieren en cierta medida en su naturaleza química, y la estructura fina de las células secretoras debe corresponder a las características de biosíntesis de cada una de ellas. No obstante, en ocasiones se pueden observar en la adenohipófisis formas transicionales de células glandulares capaces de producir varias hormonas. Existe evidencia de que el tipo de células glandulares de la adenohipófisis no siempre está determinado genéticamente.
Bajo el diafragma de la silla turca se encuentra la parte en forma de embudo del lóbulo anterior. Abraza el tallo de la glándula pituitaria y contacta con el tubérculo gris. Esta parte de la adenohipófisis se caracteriza por la presencia de células epiteliales y un abundante riego sanguíneo. Además, presenta actividad hormonal.
La parte intermedia (media) de la glándula pituitaria consta de varias capas de grandes células basófilas secretoras activas.
La hipófisis desempeña diversas funciones a través de sus hormonas. Su lóbulo anterior produce hormonas adrenocorticotrópicas (ACTH), estimulante de la tiroides (TSH), folículo estimulante (FSH), luteinizante (LH) y lipotrópicas, así como hormona del crecimiento somatotrópica (STO) y prolactina. En el lóbulo intermedio se sintetiza la hormona estimulante de los melanocitos (MSH), y en el lóbulo posterior se acumulan vasopresina y oxitocina.
ACTH
Las hormonas hipofisarias son un grupo de hormonas proteicas, peptídicas y glucoproteínas. De las hormonas de la hipófisis anterior, la ACTH es la que mejor se ha estudiado. Es producida por células basófilas. Su principal función fisiológica es estimular la biosíntesis y secreción de hormonas esteroides por la corteza suprarrenal. La ACTH también exhibe actividad lipotrópica y estimulante de melanocitos. En 1953, se aisló en forma pura. Posteriormente, se estableció su estructura química, compuesta por 39 residuos de aminoácidos en humanos y varios mamíferos. La ACTH no tiene especificidad de especie. Actualmente, se ha llevado a cabo la síntesis química tanto de la propia hormona como de varios fragmentos de su molécula, más activos que las hormonas naturales. La estructura de la hormona consta de dos secciones de la cadena peptídica: una asegura la detección y la unión de la ACTH al receptor, y la otra proporciona un efecto biológico. Al parecer, se une al receptor de ACTH mediante la interacción de las cargas eléctricas de la hormona y el receptor. El papel del efector biológico de la ACTH lo desempeña el fragmento de la molécula 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
La actividad melanocítica estimulante de la ACTH se debe a la presencia en la molécula de la región N-terminal, compuesta por 13 residuos de aminoácidos y que repite la estructura de la hormona alfa estimulante de los melanocitos. Esta región también contiene un heptapéptido presente en otras hormonas hipofisarias y que posee actividades adrenocorticotrópicas, melanocíticas y lipotrópicas.
Se considera que el momento clave en la acción de la ACTH es la activación de la enzima proteína quinasa en el citoplasma con la participación del AMPc. La proteína quinasa fosforilada activa la enzima esterasa, que convierte los ésteres de colesterol en una sustancia libre en las gotas de grasa. La proteína sintetizada en el citoplasma como resultado de la fosforilación de los ribosomas estimula la unión del colesterol libre al citocromo P-450 y su transferencia desde las gotas lipídicas a la mitocondria, donde se encuentran todas las enzimas que aseguran la conversión del colesterol en corticosteroides.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Hormona estimulante de la tiroides
La TSH (tirotropina) es el principal regulador del desarrollo y funcionamiento de la glándula tiroides, así como de los procesos de síntesis y secreción de hormonas tiroideas. Esta proteína compleja, la glicoproteína, consta de subunidades alfa y beta. La estructura de la primera subunidad coincide con la de la subunidad alfa de la hormona luteinizante. Además, es muy similar en diferentes especies animales. La secuencia de aminoácidos de la subunidad beta de la TSH humana se ha descifrado y consta de 119 aminoácidos. Cabe destacar que las subunidades beta de la TSH humana y bovina son similares en muchos aspectos. Las propiedades biológicas y la naturaleza de la actividad biológica de las hormonas glicoproteicas están determinadas por la subunidad beta. Esta también asegura la interacción de la hormona con receptores en diversos órganos diana. Sin embargo, en la mayoría de los animales, la subunidad beta exhibe actividad específica solo tras combinarse con la subunidad alfa, que actúa como un activador de la hormona. Esta última, con igual probabilidad, induce actividades luteinizantes, foliculoestimulantes y tirotrópicas, determinadas por las propiedades de la subunidad beta. La similitud encontrada permite concluir que estas hormonas surgieron en el proceso evolutivo a partir de un precursor común; la subunidad beta también determina sus propiedades inmunológicas. Se asume que la subunidad alfa protege a la subunidad beta de la acción de las enzimas proteolíticas y facilita su transporte desde la hipófisis hasta los órganos diana periféricos.
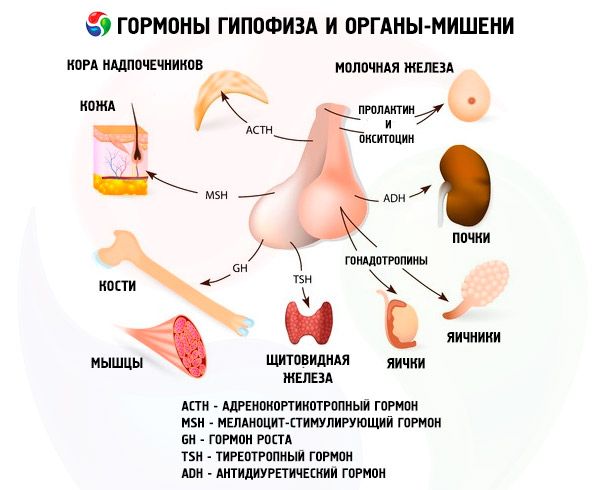
Hormonas gonadotrópicas
Las gonadotropinas están presentes en el organismo en forma de LH y FSH. Su función se limita generalmente a asegurar los procesos reproductivos en ambos sexos. Al igual que la TSH, son proteínas complejas: glicoproteínas. La FSH induce la maduración de los folículos en los ovarios femeninos y estimula la espermatogénesis en los masculinos. La LH provoca la ruptura de los folículos en las mujeres con la formación del cuerpo lúteo y estimula la secreción de estrógenos y progesterona. En los masculinos, esta misma hormona acelera el desarrollo del tejido intersticial y la secreción de andrógenos. Los efectos de las gonadotropinas son interdependientes y ocurren de forma sincrónica.
La dinámica de la secreción de gonadotropinas en las mujeres cambia durante el ciclo menstrual y se ha estudiado con suficiente detalle. En la fase preovulatoria (folicular) del ciclo, el contenido de LH es bastante bajo, mientras que el de FSH aumenta. A medida que el folículo madura, aumenta la secreción de estradiol, lo que promueve una mayor producción de gonadotropinas por la hipófisis y la aparición de ciclos de LH y FSH; es decir, los esteroides sexuales estimulan la secreción de gonadotropinas.
Actualmente, se ha determinado la estructura de la LH. Al igual que la TSH, consta de dos subunidades: a y b. La estructura de la subunidad alfa de la LH en diferentes especies animales coincide en gran medida y se corresponde con la estructura de la subunidad alfa de la TSH.
La estructura de la subunidad beta de la LH difiere significativamente de la de la subunidad beta de la TSH, aunque presenta cuatro secciones idénticas de la cadena peptídica, compuestas por 4-5 residuos de aminoácidos. En la TSH, se localizan en las posiciones 27-31, 51-54, 65-68 y 78-83. Dado que la subunidad beta de la LH y la TSH determina la actividad biológica específica de las hormonas, se puede asumir que las secciones homólogas en la estructura de la LH y la TSH deberían asegurar la conexión de las subunidades beta con la subunidad alfa, y que las secciones con diferente estructura deberían ser responsables de la especificidad de la actividad biológica de las hormonas.
La LH nativa es muy estable a la acción de las enzimas proteolíticas, pero la subunidad beta es rápidamente escindida por la quimotripsina, y la subunidad a es difícil de hidrolizar por la enzima, es decir, cumple un papel protector, impidiendo que la quimotripsina acceda a los enlaces peptídicos.
En cuanto a la estructura química de la FSH, los investigadores aún no han obtenido resultados definitivos. Al igual que la LH, la FSH consta de dos subunidades, pero la subunidad beta de la FSH difiere de la subunidad beta de la LH.
Prolactina
Otra hormona, la prolactina (hormona lactogénica), desempeña un papel activo en los procesos reproductivos. Sus principales propiedades fisiológicas en mamíferos se manifiestan en la estimulación del desarrollo de las glándulas mamarias y la lactancia, así como del crecimiento de las glándulas sebáceas y los órganos internos. Promueve la manifestación del efecto de los esteroides sobre las características sexuales secundarias en los machos, estimula la actividad secretora del cuerpo lúteo en ratones y ratas, y participa en la regulación del metabolismo de las grasas. En los últimos años, se ha prestado mucha atención a la prolactina como reguladora del comportamiento maternal; esta polifuncionalidad se explica por su desarrollo evolutivo. Es una de las hormonas hipofisarias más antiguas y se encuentra incluso en anfibios. Actualmente, la estructura de la prolactina en algunas especies de mamíferos se ha descifrado por completo. Sin embargo, hasta hace poco, los científicos expresaban dudas sobre la existencia de dicha hormona en humanos. Muchos creían que su función la desempeñaba la hormona del crecimiento. Ahora se ha obtenido evidencia convincente de la presencia de prolactina en humanos y se ha descifrado parcialmente su estructura. Los receptores de prolactina se unen activamente a la hormona del crecimiento y al lactógeno placentario, lo que indica un mecanismo de acción común de las tres hormonas.
Somatotropina
La hormona del crecimiento, somatotropina, tiene un espectro de acción aún más amplio que la prolactina. Al igual que esta, es producida por las células acidófilas de la adenohipófisis. La STH estimula el crecimiento esquelético, activa la biosíntesis de proteínas, tiene un efecto movilizador de grasa y promueve el aumento de masa corporal. Además, coordina los procesos metabólicos.
La participación de la hormona en este último se confirma por el hecho de un fuerte aumento de su secreción por la glándula pituitaria, por ejemplo, cuando disminuye el nivel de azúcar en sangre.
La estructura química de esta hormona humana está completamente establecida: 191 residuos de aminoácidos. Su estructura primaria es similar a la de la somatomamotropina coriónica o el lactógeno placentario. Estos datos indican una proximidad evolutiva significativa entre ambas hormonas, aunque presentan diferencias en su actividad biológica.
Es necesario enfatizar la alta especificidad de especie de la hormona en cuestión; por ejemplo, la STH de origen animal es inactiva en humanos. Esto se explica tanto por la reacción entre los receptores de STH de humanos y animales, como por la estructura de la propia hormona. Actualmente, se están realizando estudios para identificar centros activos en la compleja estructura de la STH que exhiban actividad biológica. Se estudian fragmentos individuales de la molécula que exhiben otras propiedades. Por ejemplo, después de la hidrólisis de la STH humana por pepsina, se aisló un péptido que consta de 14 residuos de aminoácidos y que corresponde a la sección de la molécula 31-44. No tuvo un efecto de crecimiento, pero superó significativamente a la hormona nativa en actividad lipotrópica. La hormona de crecimiento humana, a diferencia de la hormona similar de los animales, tiene una actividad lactogénica significativa.
La adenohipófisis sintetiza numerosas sustancias peptídicas y proteicas que tienen un efecto movilizador de grasa, y las hormonas hipofisarias trópicas (ACTH, STH, TSH, entre otras) tienen un efecto lipotrópico. En los últimos años, se ha prestado especial atención a las hormonas beta-lipotrópicas y y-lipotrópicas (LPG). Las propiedades biológicas de la beta-LPG se han estudiado con gran detalle; además de su actividad lipotrópica, también posee un efecto estimulante de los melanocitos, estimulante de la corticotropina e hipocalcemiante, además de producir un efecto similar a la insulina.
Actualmente, se ha descifrado la estructura primaria del LPG ovino (90 residuos de aminoácidos), hormonas lipotrópicas de cerdos y ganado vacuno. Esta hormona tiene especificidad de especie, aunque la estructura de la región central del beta-LPG es la misma en diferentes especies. Esto determina sus propiedades biológicas. Uno de los fragmentos de esta región se encuentra en la estructura de alfa-MSH, beta-MSH, ACTH y beta-LPG. Se sugiere que estas hormonas surgieron del mismo precursor en el proceso evolutivo. El γ-LPG tiene una actividad lipotrópica más débil que el beta-LPG.
Hormona estimulante de los melanocitos
Esta hormona, sintetizada en el lóbulo intermedio de la hipófisis, estimula la biosíntesis de melanina, un pigmento de la piel. En su función biológica, promueve el aumento del tamaño y la cantidad de melanocitos (células pigmentarias) en la piel de los anfibios. Estas propiedades de la MSH se utilizan en las pruebas biológicas de la hormona. Existen dos tipos de hormona: alfa-MSH y beta-MSH. Se ha demostrado que la alfa-MSH no tiene especificidad de especie y presenta la misma estructura química en todos los mamíferos. Su molécula es una cadena peptídica de 13 aminoácidos. La beta-MSH, por el contrario, sí tiene especificidad de especie y su estructura difiere entre los distintos animales. En la mayoría de los mamíferos, la molécula de beta-MSH consta de 18 aminoácidos, y solo en los humanos se extiende desde el extremo aminoacídico por cuatro aminoácidos. Cabe destacar que la alfa-MSH posee cierta actividad adrenocorticotrópica, y su efecto sobre el comportamiento de animales y humanos ha sido comprobado.
Oxitocina y vasopresina
La vasopresina y la oxitocina, sintetizadas en el hipotálamo, se acumulan en el lóbulo posterior de la hipófisis: la vasopresina en las neuronas del núcleo supraóptico y la oxitocina en el núcleo paraventricular. Posteriormente, se transfieren a la hipófisis. Cabe destacar que el precursor de la hormona vasopresina se sintetiza primero en el hipotálamo. Simultáneamente, se producen allí las proteínas neurofisinas tipos 1 y 2. La primera se une a la oxitocina y la segunda a la vasopresina. Estos complejos migran en forma de gránulos neurosecretores en el citoplasma a lo largo del axón y alcanzan el lóbulo posterior de la hipófisis, donde las fibras nerviosas terminan en la pared vascular y el contenido de los gránulos pasa a la sangre. La vasopresina y la oxitocina son las primeras hormonas hipofisarias con una secuencia de aminoácidos completamente establecida. En su estructura química, son nonapéptidos con un puente disulfuro.
Las hormonas en cuestión producen diversos efectos biológicos: estimulan el transporte de agua y sales a través de las membranas, tienen un efecto vasopresor, potencian las contracciones de la musculatura lisa del útero durante el parto y aumentan la secreción de las glándulas mamarias. Cabe destacar que la vasopresina tiene una mayor actividad antidiurética que la oxitocina, mientras que esta última tiene un efecto más potente sobre el útero y la glándula mamaria. El principal regulador de la secreción de vasopresina es el consumo de agua; en los túbulos renales, se une a receptores en las membranas citoplasmáticas, con la consiguiente activación de la enzima adenilato ciclasa. Diferentes partes de la molécula son responsables de la unión de la hormona al receptor y del efecto biológico.
La glándula pituitaria, conectada a través del hipotálamo con todo el sistema nervioso, integra el sistema endocrino en un todo funcional, contribuyendo a garantizar la constancia del ambiente interno del cuerpo (homeostasis). Dentro del sistema endocrino, la regulación homeostática se basa en el principio de retroalimentación entre el lóbulo anterior de la pituitaria y las glándulas diana (tiroides, corteza suprarrenal, gónadas). El exceso de la hormona producida por la glándula diana inhibe, y su deficiencia estimula, la secreción y liberación de la hormona trópica correspondiente. El hipotálamo forma parte del sistema de retroalimentación. En él se ubican las zonas receptoras sensibles a las hormonas de las glándulas diana. Al unirse específicamente a las hormonas que circulan en la sangre y modificar su respuesta en función de la concentración hormonal, los receptores hipotalámicos transmiten su efecto a los centros hipotalámicos correspondientes, que coordinan el trabajo de la adenohipófisis, liberando hormonas adenohipofisiotrópicas hipotalámicas. Por tanto, el hipotálamo debe considerarse como un cerebro neuroendocrino.